
感覺系統/視覺顏色感知
人類(以及猴子和猩猩等靈長類動物)在哺乳動物中擁有最佳的顏色感知能力[1]。因此,顏色在各種各樣的方面發揮重要作用並非巧合。例如,顏色對於區分和識別物體、表面、自然風景甚至面部非常有用[2],[3]。顏色也是非語言交流的重要工具,包括情緒的表達[4]。
幾十年來,找到顏色物理特性與其感知特性之間聯絡一直是一個挑戰。通常,這些研究採用兩種不同的方法:顏色引起的行為反應(也稱為心理物理學)和它引起的實際生理反應[5]。
在這裡,我們將只關注後者。顏色視覺生理基礎的研究在 20 世紀下半葉之前幾乎一無所知,從 1950 年開始緩慢而穩定地取得進展。在許多領域取得了重要進展,特別是在受體水平。由於分子生物學方法,以前未知的關於錐體色素遺傳基礎的細節得以揭示。此外,越來越多的皮質區域受到視覺刺激的影響,儘管顏色感知與受體之外的波長依賴性生理活動的相關性並不容易辨別[6]。
在本章中,我們將解釋顏色感知不同過程的基本原理,從眼球中的視網膜到大腦中的視覺皮層。有關解剖學細節,請參閱本華夏公益教科書的“視覺系統解剖學”部分。
人類可以識別的所有顏色都可以透過三種主要(基本)顏色的混合來產生。受這種顏色混合理念的啟發,人們提出顏色由三類感測器支援,每類感測器對可見光譜的不同部分具有最大敏感度[1]。1853 年,人們首次明確提出,正常顏色匹配存在三個自由度[7]。這後來在 1886 年得到證實[8](與最近的研究結果非常接近[9],[10])。
這些提議的顏色感測器實際上就是所謂的錐體細胞(注意:在本章中,我們將只討論錐體細胞。棒體細胞只在弱光條件下對視覺有貢獻。儘管已知它們會影響顏色感知,但其影響非常小,可以忽略不計。)[11]。錐體細胞是視網膜中發現的兩種光感受器細胞型別之一,其中大量集中在中央凹處。下表列出了三種類型的錐體細胞。它們透過不同的視紫紅質色素型別區分。它們相應的吸收曲線顯示在下圖中。
| 名稱 | 對顏色的更高敏感度 | 吸收曲線峰值 [nm] |
|---|---|---|
| S, SWS, B | 藍色 | 420 |
| M, MWS, G | 綠色 | 530 |
| L, LWS, R | 紅色 | 560 |
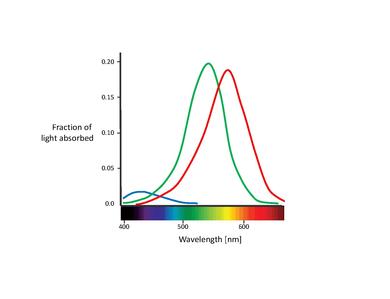
雖然對於不同錐體細胞型別的命名尚未達成共識,但最常用的名稱是指它們的行動光譜峰值或它們本身對的顏色敏感度(紅色、綠色、藍色)[6]。在這篇文字中,我們將使用 S-M-L 命名法(分別代表短、中、長波長),因為這些名稱更具描述性。“藍綠紅”命名法有些誤導,因為所有型別的錐體細胞都對很大範圍的波長敏感。
三種錐體細胞型別的一個重要特徵是它們在視網膜中的相對分佈。事實證明,S 錐體細胞在整個視網膜中的濃度相對較低,在中央凹的最中心區域完全不存在。實際上,它們之間的間距太大,無法在空間視覺中發揮重要作用,儘管它們能夠介導弱邊界感知[12]。中央凹主要由 L 錐體細胞和 M 錐體細胞組成。後兩種型別的比例通常以比率衡量。L/M 比率報告了不同的值,範圍從 0.67[13]到 2[14],後者是最被接受的值。L 錐體細胞幾乎總是比 M 錐體細胞數量多,其原因尚不清楚。令人驚訝的是,相對錐體細胞比例幾乎對顏色視覺沒有顯著影響。這清楚地表明,大腦具有可塑性,能夠從它接收到的任何錐體細胞訊號中提取意義[15],[16]。
還需要注意的是 L 錐體細胞和 M 錐體細胞吸收光譜的重疊。雖然 S 錐體細胞吸收光譜是明顯分開的,但 L 錐體細胞和 M 錐體細胞峰值之間只有約 30 nm 的距離,它們的光譜曲線也存在顯著的重疊。這導致這兩類錐體細胞的感光量高度相關。這可以用這樣一個事實來解釋:為了在中央凹中心實現儘可能高的銳度,視覺系統平等地對待 L 錐體細胞和 M 錐體細胞,而不考慮它們的光譜吸收。因此,任何差異都會導致亮度訊號惡化[17]。換句話說,L 錐體細胞和 M 錐體細胞光譜之間的微小分離可以解釋為高對比度顏色視覺和高銳度亮度視覺需求之間的折衷。這與中央凹中心部分沒有 S 錐體細胞相一致,因為中央凹中心部分的視覺銳度最高。此外,L 錐體細胞和 M 錐體細胞吸收光譜之間的緊密間距也可以用它們的遺傳起源來解釋。據推測,這兩類錐體細胞都是從一個共同的祖先最近(大約 3500 萬年前)進化而來的,而 S 錐體細胞可能在更早之前就從祖先受體中分離出來[11]。
三種不同型別錐體細胞的光譜吸收函式是人類顏色視覺的標誌。該理論解決了一個長期存在的問題:雖然我們能看到數百萬種不同的顏色(人類可以區分 700 萬到 1000 萬種不同的顏色[5]),但我們的視網膜 simply does not have enough space to accommodate an individual detector for every color at every retinal location.
從視網膜傳遞到更高層次的訊號不是受體訊號的簡單逐點表示,而是受體訊號的複雜組合。本節的目的是簡要介紹部分資訊傳遞的路徑。
一旦視網膜上的光學影像被轉換為光感受器中的化學和電訊號,幅度調製訊號就會在神經節細胞和更高層次轉換為頻率調製表示。在這些神經細胞中,訊號的幅度用細胞每秒發出的電壓尖峰數量表示,而不是用細胞膜上的電壓差表示。為了解釋和表示這些細胞的生理特性,我們會發現感受野的概念非常有用。
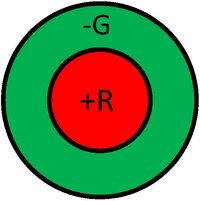
感受野是視覺區域中給定細胞響應區域的圖形表示。此外,響應的性質通常在感受野的不同區域中表示出來。例如,我們可以將光感受器的感受野視為一個小的圓形區域,代表該特定受體在視覺區域的靈敏度的大小和位置。下圖顯示了神經節細胞的示例性感受野,通常以中心-周圍拮抗的形式。圖中左側感受野說明了正中心響應(稱為中心-ON)。這種響應通常是由單個錐體產生的正輸入以及由幾個相鄰錐體產生的負響應共同生成的。因此,這種神經節細胞的響應將由來自不同錐體的正負輸入組成。這樣一來,細胞不僅對光點做出反應,而且充當邊緣(更準確地說,是斑點)探測器。類比於計算機視覺術語,我們可以將神經節細胞的響應視為對邊緣探測器核卷積後的輸出。圖中右側感受野說明了負中心響應(稱為中心-OFF),其可能性相同。通常,中心-ON和中心-OFF細胞將出現在同一空間位置,由相同的光感受器供給,從而導致增強的動態範圍。
下圖顯示了除了空間拮抗之外,神經節細胞還可以具有光譜對抗性。例如,下圖左側說明了紅-綠對抗響應,其中中心由來自L錐體的正輸入供給,周圍由來自M錐體的負輸入供給。另一方面,下圖右側說明了這種細胞的中心-OFF版本。因此,在視覺資訊甚至離開視網膜之前,處理就已經發生了,對顏色外觀產生了深刻的影響。還有其他型別和品種的神經節細胞響應,但它們都共享這些基本概念。



在通往初級視覺皮層的路上,神經節細胞的軸突聚集起來形成視神經,它投射到丘腦的背側膝狀核(LGN)。視神經中的編碼非常高效,使神經纖維的數量降至最低(受視神經大小的限制),從而也使視網膜盲點的尺寸儘可能小(約 5° 寬 x 7° 高)。此外,所呈現的神經節細胞對均勻照度將沒有反應,因為正負區域是平衡的。換句話說,傳輸的訊號是不相關的。例如,來自自然場景相鄰部分的資訊在空間上高度相關,因此高度可預測[18]。相鄰視網膜神經節細胞之間的側抑制最大程度地減少了這種空間相關性,從而提高了效率。我們可以將其視為在視網膜中進行的一種影像壓縮過程。
鑑於L-和M-錐體吸收光譜的重疊,它們的訊號也高度相關。在這種情況下,透過組合錐體訊號來最大程度地減少這種相關性,可以提高編碼效率。我們可以使用主成分分析 (PCA) 更容易地理解這一點。PCA 是一種統計方法,用於透過將原始變數轉換為一組新的變數(主成分 (PC))來降低給定變數集的維數。第一個 PC 解釋了原始變數中總方差的最大量,第二個 PC 解釋了第一個成分未解釋的最大方差,依此類推。此外,PC 在引數空間中線性無關且彼此正交。PCA 的主要優點是,只有幾個最強的 PC 就足以覆蓋系統變異性的絕大多數[19]。這種方案已應用於錐體吸收函式[20],甚至應用於自然發生的頻譜[21],[22]。在錐體激發空間中發現的 PC 是 1) 亮度軸,其中 L- 和 M- 錐體訊號相加 (L+M),2) L- 和 M- 錐體訊號的差 (L-M),以及 3) 色彩軸,其中 S- 錐體訊號與 L- 和 M- 錐體訊號的總和之差 (S-(L+M))。這些從數學/計算方法匯出的通道與在電生理實驗中發現的三個視網膜-膝狀核通道相吻合[23],[24]。使用這些機制,視覺冗餘資訊在視網膜中被消除。
實際上,有三個資訊通道將資訊從視網膜透過神經節細胞傳遞到 LGN。它們不僅在色度特性上不同,而且在解剖學基礎上也不同。這些通道對基本顏色任務(如檢測和辨別)構成重要限制。
在第一個通道中,L- 和 M- 錐體的輸出協同傳遞到彌散雙極細胞,然後傳遞到 LGN 的大細胞層 (M-) 中的細胞(不要與視網膜的 M- 錐體混淆)[24]。M- 細胞的感受野由中心和周圍組成,它們在空間上是拮抗的。M- 細胞對亮度刺激具有高對比度敏感性,但它們在 L-M 對抗輸入的某些組合下沒有反應[25]。然而,由於不同 M- 細胞的零點略有不同,總體響應實際上永遠不會真正為零。這種特性實際上被傳遞到以 M- 細胞輸入為主的皮層區域[26]。
小細胞通路 (P-) 起源於 L- 或 M- 錐體到矮胖雙極細胞的單個輸出。它們為視網膜 P- 細胞提供輸入[11]。在中央凹,P- 細胞的感受野中心由單個 L- 或 M- 錐體形成。P- 細胞感受野周圍的結構仍在爭論中。然而,最被接受的理論指出,周圍由特定型別的錐體組成,導致亮度刺激的空間拮抗感受野[27]。小細胞層約佔視網膜到 LGN 總投射的 80%[28]。
最後,最近發現的細小細胞通路 (K-) 主要攜帶來自 S- 錐體的訊號[29]。這種型別錐體的群體投射到特殊的雙極細胞,而這些雙極細胞反過來為特定的小型神經節細胞提供輸入。這些通常不是空間拮抗的。小型神經節細胞的軸突投射到 LGN 的薄層(緊鄰小細胞層)[30]。
雖然神經節細胞確實終止於 LGN(與 LGN 細胞形成突觸),但神經節細胞和 LGN 細胞之間似乎存在一對一對應關係。LGN 似乎充當訊號的繼電站。然而,它可能在視覺功能中發揮作用,因為皮層存在神經投射回 LGN,可以作為某種型別的切換或適應反饋機制。LGN 細胞的軸突投射到枕葉的視覺區一 (V1)。
在皮層中,來自大細胞、小細胞和細小細胞通路的投射最終到達初級視覺皮層的不同層。大細胞纖維主要支配 4Cα 層和 6 層。小細胞神經元主要投射到 4Cβ、4A 層和 6 層。細小細胞神經元終止於 1、2 和 3 層的細胞色素氧化酶 (CO-) 富集斑點[31]。
一旦進入視覺皮層,視覺資訊的編碼變得更加複雜。與各種光感受器的輸出組合並比較以產生神經節細胞響應的方式相同,各種 LGN 細胞的輸出被比較並組合起來以產生皮層響應。隨著訊號在皮層處理鏈中進一步向上推進,這個過程會重複進行,複雜程度迅速增加,以至於感受野開始失去意義。然而,一些功能和過程已在視覺皮層的特定區域中被識別和研究。
在 V1 區域(紋狀皮層),雙拮抗神經元(其感受野在色度和空間上都與其單個感受野的興奮/抑制區域相反)比較視覺空間中的顏色訊號[32]。它們佔 V1 中細胞的 5% 到 10%。它們粗糙的大小和較小的比例與顏色視覺的空間解析度差相匹配[1]。此外,它們對移動刺激的方向不敏感(不像其他一些 V1 神經元),因此不太可能對運動感知做出貢獻[33]。然而,鑑於它們特殊的感受野結構,這類細胞是顏色對比效應的神經基礎,也是編碼顏色的有效手段[34],[35]。其他 V1 細胞對其他型別的刺激做出反應,例如定向邊緣、各種空間和時間頻率、特定空間位置以及這些特徵的組合等等。此外,我們可以找到線性組合來自 LGN 細胞的輸入的細胞,以及執行非線性組合的細胞。這些反應是支援高階視覺能力所必需的,例如顏色本身。

與 V1 相比,V2 中單個神經元の色度特性資訊要少得多。乍一看,V1 和 V2 的顏色編碼似乎沒有重大差異[36]。一個例外是出現了一類新的顏色複雜細胞[37]。因此,有人認為 V2 區域參與了色調的闡述。然而,這仍然存在很大爭議,尚未得到證實。
在 V1 中發現功能性眼優勢後,按照模組化概念,並考慮到 P、M 和 K 通路之間的解剖學分離(在第 3 節中描述),有人提出視覺皮層中應該存在一個專門的系統來分析顏色資訊[38]。V4 是歷史上最受關注的可能的大腦“顏色區域”。這是因為一項有影響力的研究聲稱 V4 包含 100% 的色調選擇性細胞[39]。然而,這一說法受到隨後多項研究的質疑,有些研究甚至報告說,只有 16% 的 V4 神經元表現出色調調節[40]。目前,最被接受的概念是 V4 不僅對顏色有貢獻,而且對形狀感知、視覺注意力和立體視覺也有貢獻。此外,最近的研究集中在其他大腦區域,試圖找到大腦的“顏色區域”,例如 TEO[41] 和 PITd[42]。這些區域之間的關係仍在爭論中。為了調和討論,有些人使用術語“後顳下(PIT)皮層”來表示包括 V4、TEO 和 PITd 的區域[1]。
如果 V1、V2 和 V4 細胞中的皮層響應已經是一項非常複雜的任務,那麼大約 30 個視覺區域網路中複雜視覺響應的複雜程度就非常巨大。圖 4 顯示了已識別的不同皮層區域(不是細胞)的連線性的一小部分[43]。
在這個階段,用簡單的術語解釋單個皮層細胞的功能變得極其困難。事實上,單個細胞的功能可能沒有意義,因為各種感知的表徵必須分佈在整個皮層中的一組細胞中。
顏色視覺適應機制
[edit | edit source]儘管研究人員一直在試圖解釋人類視覺系統中顏色訊號的處理過程,但重要的是要理解顏色感知不是一個固定過程。實際上,有各種動態機制用於根據觀看環境最佳化視覺響應。與顏色感知特別相關的是暗適應、亮適應和色度適應的機制。
暗適應
[edit | edit source]暗適應是指當照明水平降低時發生的視覺靈敏度變化。視覺系統對光照降低的反應是變得更加敏感,提高其即使在光照條件不佳的情況下也能產生有意義的視覺響應的能力[44]。
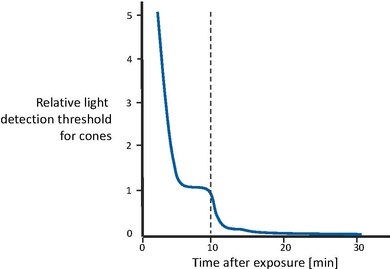
圖 5 顯示了從極高照明水平過渡到完全黑暗後視覺靈敏度的恢復[43]。首先,視錐細胞逐漸變得更加敏感,直到在幾分鐘後曲線趨於平穩。然後,在大約 10 分鐘後,視覺靈敏度大致保持恆定。此時,視杆系統恢復了足夠的靈敏度,其恢復時間更長,能夠勝過視錐細胞,因此控制了整體靈敏度。視杆細胞靈敏度也逐漸提高,直到在大約 30 分鐘後變得漸近。換句話說,視錐細胞負責最初 10 分鐘的靈敏度恢復。之後,視杆細胞的效能優於視錐細胞,在大約 30 分鐘後獲得完全靈敏度。
這只是為了儘可能好地適應黑暗光照條件而產生的幾種神經機制之一。其他一些神經機制包括著名的瞳孔反射、光色素的消耗和再生、視網膜細胞中的增益控制和其他更高級別的機制以及認知解釋等等。
亮適應
[edit | edit source]亮適應本質上是暗適應的逆過程。事實上,這兩個過程的基礎生理機制是相同的。但是,重要的是要單獨考慮它,因為它的視覺特性有所不同。

亮適應發生在照明水平提高時。因此,視覺系統必須變得不那麼敏感,以便在有大量可見光可用時產生有用的感知。視覺系統用於產生感知的訊號具有有限的輸出動態範圍。但是,現實世界中的照明水平至少覆蓋了 10 個數量級的變化。幸運的是,我們很少需要同時檢視整個照明範圍。
在高光照水平下,適應是透過光色素漂白實現的。這會調整受體中的光子捕獲,並保護視錐細胞響應在明亮背景下不被飽和。亮適應的機制主要發生在視網膜內[45]。事實上,增益變化在很大程度上是視錐細胞特異性的,並且適應會將訊號池化到不超過單個視錐細胞直徑的區域[46],[47]。這表明亮適應的定位可能早在受體階段就開始了。然而,似乎存在不止一個靈敏度縮放位點。有些增益變化非常快,而另一些則需要幾秒甚至幾分鐘才能穩定[48]。通常,亮適應需要大約 5 分鐘(比暗適應快六倍)。這可能表明了受體後位點的影響。
圖 6 顯示了亮適應的示例[43]。如果我們使用單個響應函式將大範圍的強度對映到視覺系統的輸出,那麼對於給定的場景,我們只能使用非常小的範圍。很明顯,使用這種響應函式,任何給定場景的感知對比度將受到限制,並且由於信噪比問題,對變化的視覺靈敏度會嚴重降低。這種情況由虛線表示。另一方面,實線代表視覺響應的曲線族。這些曲線將給定場景中的有用照明範圍對映到視覺輸出的完整動態範圍,從而為每種情況產生最佳的視覺感知。亮適應可以看作是將視覺響應曲線沿著照明水平軸滑動,直到達到給定觀看條件下的最佳水平。
色彩適應的概念通常是指三種錐體光譜響應曲線高度的變化。這種調整的出現是因為光適應在每種錐體型別中獨立發生。該假設的具體公式被稱為 *馮·克萊斯適應*。該假設指出,適應反應分別在三種錐體型別中發生,並且等效於將它們固定的光譜敏感度乘以一個比例常數[49]。如果比例權重(也稱為 *馮·克萊斯係數*)與每種錐體型別的光吸收成反比(即,較低的光吸收需要較大的係數),那麼馮·克萊斯縮放將保持每種錐體類別內的平均響應恆定。這為保持物體在照明變化的情況下感知到的顏色提供了一種簡單而強大的機制。在多種不同的條件下,馮·克萊斯縮放很好地解釋了光適應對顏色敏感度和外觀的影響[50],[51]。
觀察不同照明條件下白色物體是最容易理解色彩適應的方式。例如,考慮在日光、熒光燈和白熾燈下觀察一張紙。日光含有比熒光燈更多的短波能量,而白熾燈含有比熒光燈更多的長波能量。然而,儘管照明條件不同,紙張在三種光源下都近似保持其白色外觀。這是因為 S 錐體系統在日光下變得相對不那麼敏感(為了補償額外的短波能量),而 L 錐體系統在白熾燈下變得相對不那麼敏感(為了補償額外的長波能量)[43]。
- ↑ a b c d Conway, Bevil R (2009). "顏色視覺、錐體和皮層中的顏色編碼". 神經科學家. 15: 274–290.
- ↑ Russell, Richard and Sinha, Pawan} (2007). "現實世界中的面部識別:表面反射屬性的重要性". 知覺. 36 (9).
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Gegenfurtner, Karl R and Rieger, Jochem (2000). "顏色對自然場景識別的感官和認知貢獻". 當代生物學. 10 (13): 805–808.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Changizi, Mark A and Zhang, Qiong and Shimojo, Shinsuke (2006). "裸露的皮膚、血液和靈長類顏色視覺的進化". 生物學快報. 2 (2): 217–221.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ a b Beretta, Giordano (2000). 理解顏色. 惠普公司.
- ↑ a b Boynton, Robert M (1988). "顏色視覺". 心理學年度回顧. 39 (1): 69–100.
- ↑ Grassmann, Hermann (1853). "關於顏色混合理論". 物理學年鑑. 165 (5): 69–84.
- ↑ Konig, Arthur and Dieterici, Conrad (1886). "基本感覺及其在光譜中的強度分佈". 普魯士皇家科學院.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Smith, Vivianne C and Pokorny, Joel (1975). "中央凹錐體光感受器在 400 到 500 奈米之間的光譜敏感度". 視覺研究. 15 (2): 161–171.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ↑ Vos, JJ 和 Walraven, PL (1971)。“關於中央凹視錐細胞原色的推導”。視覺研究。11 (8): 799–818.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ a b c Gegenfurtner, Karl R 和 Kiper, Daniel C (2003)。“色覺”。神經科學。26 (1): 181.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Kaiser, Peter K 和 Boynton, Robert M (1985)。“藍色機制在波長辨別中的作用”。視覺研究。125 (4): 523–529.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Paulus, Walter 和 Kroger-Paulus, Angelika (1983)。“視網膜顏色編碼的新概念”。視覺研究。23 (5): 529–540.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Nerger, Janice L 和 Cicerone, Carol M (1992)。“人類旁中央凹視網膜中 L 視錐細胞與 M 視錐細胞的比例”。視覺研究。32 (5): 879–888.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Neitz, Jay 和 Carroll, Joseph 和 Yamauchi, Yasuki 和 Neitz, Maureen 和 Williams, David R (2002)。“色覺由一種可塑的神經機制介導,這種機制在成年人中是可調節的”。神經元。35 (4): 783–792.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Jacobs, Gerald H 和 Williams, Gary A 和 Cahill, Hugh 和 Nathans, Jeremy (2007)。“在表達人類視錐細胞光色素的轉基因小鼠中出現的新的色覺”。科學。315 (5819): 1723–1725.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Osorio, D 和 Ruderman, DL 和 Cronin, TW (1998)。“估計靈長類動物視網膜編碼的亮度訊號中由紅視錐細胞和綠視錐細胞對自然影像進行取樣而產生的誤差”。JOSA A。15 (1): 16–22.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Kersten, Daniel (1987)。“自然影像的可預測性和冗餘”。JOSA A。4 (112): 2395–2400.
- ↑ Jolliffe, I. T. (2002)。主成分分析。施普林格出版社.
- ↑ Buchsbaum, Gershon 和 Gottschalk, A (1983)。“三色視覺、對立色編碼和視網膜中的最佳顏色資訊傳輸”。倫敦皇家學會會刊。B輯。生物科學。220 (1218): 89–113.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Zaidi, Qasim (1997)。“L 視錐細胞和 M 視錐細胞訊號的去相關”。JOSA A。14 (12): 3430–3431.
- ↑ Ruderman, Daniel L 和 Cronin, Thomas W 和 Chiao, Chuan-Chin (1998)。“視錐細胞對自然影像的響應統計:對視覺編碼的意義”。JOSA A。15 (8): 2036–2045.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Lee, BB 和 Martin, PR 和 Valberg, A (1998). "異色閃爍光度計的生理基礎,在獼猴視網膜神經節細胞中得到證實". 生理學雜誌. 404 (1): 323–347.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ a b Derrington, Andrew M 和 Krauskopf, John 和 Lennie, Peter (1984). "獼猴外側膝狀體核中的色覺機制". 生理學雜誌. 357 (1): 241–265.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Shapley, Robert (1990). "視覺敏感度和並行的視網膜皮層通路". 心理學年度評論. 41 (1): 635--658.
- ↑ Dobkins, Karen R 和 Thiele, Alex 和 Albright, Thomas D (2000). "人類和獼猴紅綠等亮度點的比較:不同物種間 L:M 視錐比例的證據". JOSA A. 17 (3): 545–556.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Martin, Paul R 和 Lee, Barry B 和 White, Andrew JR 和 Solomon, Samuel G 和 Ruttiger, Lukas (2001). "靈長類動物外周視網膜神經節細胞的色覺敏感性". 自然. 410 (6831): 933–936.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Perry, VH 和 Oehler, R 和 Cowey, A (1984). "投射到獼猴背側外側膝狀體核的視網膜神經節細胞". 神經科學. 12 (4): 1101--1123.
{{cite journal}}: 引文格式有誤:未知引數忽略:|month=(help)CS1 maint: 多個名字:作者列表 (link) - ↑ Casagrande, VA (1994). "通往靈長類動物 V1 區的第三條平行視覺通路". 神經科學趨勢. 17 (7): 305–310.
- ↑ Hendry, Stewart HC 和 Reid, R Clay (2000). "靈長類視覺中的錐體通路". 神經科學年度評論. 23 (1): 127–153.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Callaway, Edward M (1998). "獼猴初級視覺皮層的區域性迴路". 神經科學年度評論. 21 (1): 47–74.
- ↑ Conway, Bevil R (2001). "清醒獼猴初級視覺皮層 (V-1) 中色覺細胞的視錐輸入的空間結構". 神經科學雜誌. 21 (8): 2768–2783.
{{cite journal}}: 引文格式有誤:未知引數忽略:|month=(help) - ↑ Horwitz, Gregory D 和 Albright, Thomas D (2005). "獼猴 V1 中色覺線性運動檢測器的稀缺". 視覺雜誌. 5 (6).
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Danilova, Marina V 和 Mollon, JD (2006). "空間分離顏色的比較". 視覺研究. 46 (6): 823–836.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Wachtler, Thomas 和 Sejnowski, Terrence J 和 Albright, Thomas D (2003). "清醒獼猴初級視覺皮層中色覺刺激的表徵". 神經元. 37 (4): 681–691.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Solomon, Samuel G 和 Lennie, Peter (2005). "視覺皮層神經元中的色覺增益控制". 神經科學雜誌. 25 (19): 4779–4792.
{{cite journal}}: CS1 maint: 多個名字:作者列表 (link) - ↑ Hubel, David H (1995). 眼、腦和視覺. 科學美國人圖書館/科學美國人書籍.
- ↑ Livingstone, Margaret S 和 Hubel, David H (1987). “心理物理學證據表明,感知形狀、顏色、運動和深度的通道是分開的”。神經科學雜誌. 7 (11): 3416–3468.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ Zeki, Semir M (1973). “獼猴前紋狀皮層的顏色編碼”。腦研究. 53 (2): 422–427.
- ↑ Conway, Bevil R 和 Tsao, Doris Y (2006). “fMRI 揭示清醒獼猴皮層的顏色結構”。大腦皮層. 16 (11): 1604–1613.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ Tootell, Roger BH、Nelissen, Koen、Vanduffel, Wim 和 Orban, Guy A (2004)。“在獼猴視覺皮層中尋找顏色‘中心’”。大腦皮層. 14 (4): 353–363.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ Conway, Bevil R、Moeller, Sebastian 和 Tsao, Doris Y (2007)。“獼猴外紋狀皮層的特化顏色模組”。560--573. 56 (3): 560–573.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ a b c d Fairchild, Mark D (2013)。顏色外觀模型. 約翰·威利父子公司.
- ↑ Webster, Michael A (1996)。“人類顏色感知及其適應”。網路:神經系統中的計算. 7 (4): 587–634.
- ↑ Shapley, Robert 和 Enroth-Cugell, Christina (1984)。“視覺適應和視網膜增益控制”。視網膜研究進展. 3: 263–346.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ Chaparro, A、Stromeyer III, CF、Chen, G 和 Kronauer, RE (1995)。“人類視錐細胞似乎在低光照條件下會適應:對紅綠檢測機制的測量”。視覺研究. 35 (22): 3103–3118.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ Macleod, Donald IA、Williams, David R 和 Makous, Walter (1992)。“由單個視錐細胞饋送的視覺非線性”。視覺研究. 32 (2): 347–363.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ Hayhoe, Mary (1991)。顏色和亮度的適應機制. 春季.
- ↑ MacAdam, DAvid L (1970)。顏色科學的來源. 麻省理工學院出版社.
- ↑ Webster, Michael A 和 Mollon, JD (1995)。“對比度適應影響的顏色恆常性”。自然. 373 (6516): 694–698.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link) - ↑ Brainard, David H 和 Wandell, Brian A (1992)。“不對稱顏色匹配:顏色外觀如何依賴於光源”。JOSA A. 9 (9): 1443–1448.
{{cite journal}}: CS1 maint: 多個名稱:作者列表 (link)