
蛋白質組學/蛋白質一級結構/選擇性剪接
本節
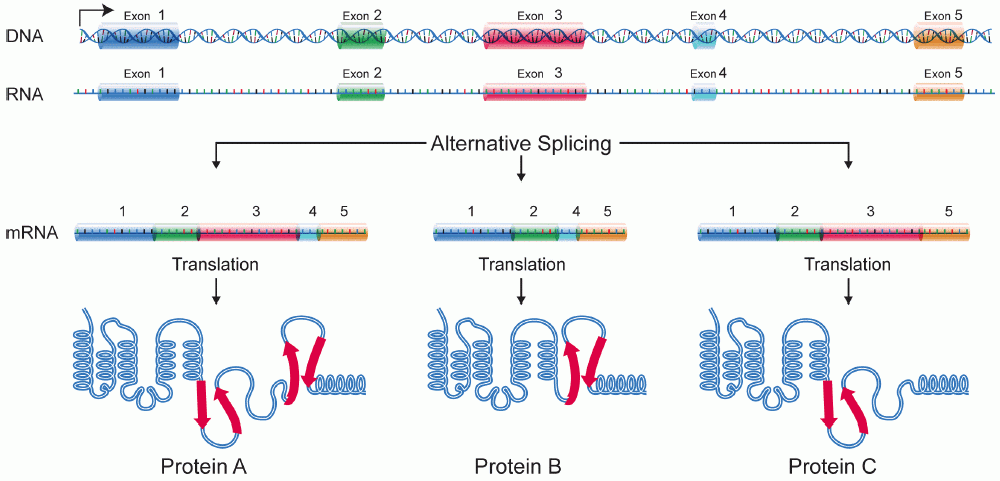
選擇性剪接是指基因的初級轉錄本被重組以產生多種成熟的mRNA轉錄本的過程。透過操縱外顯子區域,可以控制從mRNA產生的氨基酸序列,從而產生具有不同結構和功能的不同蛋白質序列。 [1] 選擇性剪接已被觀察到是產生來自單個基因的組織特異性蛋白質的機制。這允許增加細胞特化,而無需基因修飾。這是發育的一個非常關鍵的方面,因為它允許以仍然允許靈活性,和修飾的方式確定細胞命運。選擇性剪接,尤其是在神經元發育的背景下進行了研究, [2] [3] 然而,許多剪接變體已被發現,它們僅在發育期間的某些組織中表達,似乎在觸發細胞型別特異性基因表達中起著至關重要的作用。 [4]

{kind=link}
選擇性剪接是生物體中蛋白質多樣性的主要來源。據估計,人類基因組中至少 30% 的基因都經歷選擇性剪接,並且這個數字還在不斷增加。最初認為,選擇性剪接基因的數量僅佔人類蛋白質的 5%。隨著人類基因組的揭示,發現人類基因組包含不到 30,000 個基因 [5]。這可能可以解釋相對較少的基因數量和蛋白質組中大量蛋白質之間的巨大差距。
有人提出,選擇性剪接是真核生物中高階複雜性的來源 [6]。這個想法是基於這樣一個想法,即更復雜的生物體將更頻繁地選擇性剪接其基因以獲得更多可能的 mRNA 序列。然而,證據表明,不同複雜程度的生物體之間的選擇性剪接水平並不顯著。一項使用 EST(表達序列標籤 [7])進行的研究提供了相反的證據。EST 作為轉錄的 mRNA 的一部分,在檢測選擇性剪接基因方面非常有效。隨著越來越多的 EST 研究的開展,越來越明顯的是,選擇性剪接基因的數量比以前認為的要多(見上文)。
選擇性剪接與多種疾病有關。一個在選擇性剪接中起作用的疾病的例子是雷特綜合徵 (RTT)。這種疾病主要發生在女孩身上,其特徵是神經元之間連線形成障礙,或突觸 [8]患有這種疾病的女孩通常在 6-18 個月大之前發育正常 [9]。在這個年齡之後,她們開始表現出運動和語言技能下降,通常伴隨有搓手動作。 [10] 據信,MeCP2 基因會產生一種突變蛋白,該蛋白會破壞其作為某些蛋白質中選擇性剪接的共調節因子的正常功能。當這種突變版本的 MeCP2 產生時,會產生一種缺乏野生型關鍵功能的蛋白質,該蛋白質不允許調節選擇性剪接。MeCP2 最初被確定為一種甲基化依賴的轉錄抑制因子。然而,人們發現 MeCP2 具有其他功能,這是由 MeCP2 與 Y 盒結合蛋白 (YB-1,mRNP 的一個組分) 的相互作用首先闡明的。YB-1 和 MeCP2 的複合物被證明可以調節報告基因的剪接。
來自導致表型類似於雷特綜合徵的突變小鼠的微陣列資料顯示,選擇性剪接基因的數量與野生型小鼠相比存在顯著差異。選擇性剪接轉錄本與細胞分析相匹配,細胞分析表明 MeCP2 在盒式外顯子改變中發揮作用。
剪接透過剪接體機制發生,剪接體是一種由多種蛋白質和 snRNA 組成的複合物,包括 snRNA U1、U2、U3、U4、U5 和 U6 [11]。這些 snRNA 識別剪接位點,然後募集其他蛋白質來連線剪接位點。然後,這些剪接位點透過這些形成剪接體的蛋白質的相互作用被連線在一起。一旦剪接體形成,這些位點就會被切割以使正確的外顯子(或內含子)連線在一起。生物體中的剪接位點尚未得到很好的表徵。在使用 HMM 分析和其他形式的機器學習進行計算預測方面已經取得了一些進展。這將在本文後面討論。內含子的 5' 端以通常以“GU”結尾的共有序列為特徵。該區域被稱為剪接供體位點。內含子的 3' 端被稱為剪接受體位點,包含一個共有序列,該序列在內含子/外顯子邊界處具有一個 AG。在剪接機制中起作用的還有靠近內含子 3' 端的一個腺嘌呤,通常稱為分支點。在剪接過程中,U1 開始在剪接供體位點形成複合物,而 U2 將剪接體複合物的一部分募集到剪接受體位點。U4、U5 和 U6 連線這兩個位點之間的間隙,導致內含子摺疊。DNA 在剪接供體位點被消化。然後,內含子的這部分與分支點連線,形成套索。這使外顯子區域彼此靠近,允許連線。最後,剪接受體位點被切割,外顯子區域被連線起來。 [12]

{kind=link}
有 4 種常見的選擇性剪接型別,如下所示
- 選擇性啟動子選擇:不同的啟動子用於不同的剪接變體。這導致 mRNA 轉錄本的起點不同。
- 選擇性切割/聚腺苷酸化位點:基於對不同切割或聚腺苷酸化位點的識別,不同的外顯子被剪接,整個外顯子可以被跳過。導致轉錄本 3' 端的外顯子不同。
- 內含子保留:內含子用作編碼區。通常被認為是內含子的序列被保留在最終轉錄本中,該轉錄本充當翻譯的模板。
- 外顯子盒:整個外顯子可以在蛋白質中間被跳過,導致不同的轉錄本

{kind=link}
蛋白質是生物系統中結構和功能的基本單位。因此,蛋白質組學領域在現代生物學研究中變得更加重要。基因組革命,其最終導致了多個基因組的測序,產生了大量資料。蛋白質組學領域 [13] 不幸的是落後了,導致基因組資訊和可觀察到的表型之間脫節。最初,蛋白質和蛋白質依賴的途徑是單獨研究的。最近,對系統生物學的強調導致了這種方法的改變。整個細胞正在使用高通量技術進行表徵。

{kind=link}
質譜法 已成為蛋白質鑑定的金標準 [14] [15]。簡而言之,蛋白質被分解成肽段,透過多種方法之一將其懸浮到氣相中,電離,然後送入檢測器,檢測器可以確定各種肽段的質荷比。質譜法易於自動化,並可與其他形式的蛋白質分離方法結合使用,使其成為高通量分析的理想選擇。此外,可以一次從單個來源識別數千種肽段,使該技術比埃德曼降解等舊技術更適用於系統生物學。質譜法還可以用於識別已使用其他技術(如色譜法)分離的特定感興趣的蛋白質。

{kind=link}
剪接和蛋白質鑑定
[edit | edit source]質譜分析甚至埃德曼降解鑑定的一個主要缺點是,蛋白質必須在鑑定之前被消化成肽段。通常為了與鳥槍法測序一起使用 [16],蛋白質在任何形式的隔離過程完成之前就被消化。為了確認鑑定結果,搜尋蛋白質資料庫以匹配唯一肽段到完整蛋白質。這種過程因可變剪接產生的肽段之間存在大量序列同源性而變得更加複雜。 [17]。這些蛋白質雖然具有相似的初級結構,但可能具有非常不同的甚至拮抗的功能,因此從生物學的角度來看,識別它們至關重要。更重要的是,可變剪接發生的程度尚未得到很好的表徵,因此甚至不知道哪些蛋白質無法被明確識別。這些問題可能在可變剪接得到充分記錄或能夠透過計算有效地預測之前一直存在。
分析可變剪接的影響
[edit | edit source]體內分析目前是識別可變剪接的最準確方法,無論是在轉錄本水平還是在某些情況下在蛋白質水平。許多資料庫記錄了已知經歷可變剪接的蛋白質,包括可變剪接資料庫 [18] 和轉錄本多樣性資料庫 [19]。雖然這些都是很好的參考資料,但如上所述,即使是對真核生物中存在多少可變剪接的估計,也存在很大差異。因此,關於這些資料庫的完整性資訊非常少。
還採取了一些步驟來計算地預測可變剪接 [20]。通常,這些演算法將基因尋找方法與實驗資料結合在一起。剪接位點被識別並根據一致性序列對其效率進行評分。然後將序列與已知的表達序列標籤匹配以進行預測。BLAST、Spidey 和 SIM4 等工具可以適用於這些過程。大多數現代計算工具在將基因組資料與剪接位點一樣小且多變的序列進行比較時會遇到困難。假陽性和假陰性相當普遍 [21]。新的方法仍在開發中。
參考文獻
[edit | edit source]- ↑ Möröy, T 等人。 “體內可變剪接的影響:小鼠模型指明瞭方向”。 http://www.rnajournal.org/cgi/content/full/13/8/1155 於 2008 年 4 月獲得
- ↑ Goymer, P. “發育:可變剪接開啟大腦”。《自然評論神經科學》8,576(2007 年 8 月)。 http://www.nature.com/nrn/journal/v8/n8/full/nrn2200.html 於 2008 年 4 月獲得
- ↑ Azuma, N 等人。 “帶有可變剪接外顯子的 Pax6 同種型促進神經視網膜結構的發育”。《人類分子遺傳學》2005 年 14(6):735-745 http://hmg.oxfordjournals.org/cgi/content/full/14/6/735 於 2008 年 4 月獲得
- ↑ Michelle Lesimple 等人。《DNA 和細胞生物學》。2000 年 6 月 1 日,19(6): 365-376 http://www.liebertonline.com/doi/abs/10.1089/10445490050043335 於 2008 年 4 月獲得。
- ↑ 可變 RNA 剪接。ExonHit Therapeutics。 http://www.exonhit.com/index.php?page=59。於 2008 年 4 月獲得
- ↑ Brett D 等人。 可變剪接和基因組。《自然遺傳學》30,29-30(2001) http://www.nature.com/ng/journal/v30/n1/abs/ng803.html;jsessionid=BF0AED8347574D063F5E347EC693AE83 於 2008 年 4 月獲得
- ↑ http://en.wikipedia.org/wiki/Expressed_sequence_tag
- ↑ “雷特綜合徵概況”。美國國立神經疾病和中風研究所。 http://www.ninds.nih.gov/disorders/rett/detail_rett.htm#109713277 於 2008 年 4 月獲得。
- ↑ “雷特基因調節可變剪接”。《科學日報》。 http://www.sciencedaily.com/releases/2005/10/051019002531.htm 於 2008 年 4 月獲得
- ↑ Young, J 等人。 “依賴甲基化的轉錄抑制因子甲基-CpG 結合蛋白 2 對 RNA 剪接的調節”。《美國國家科學院院刊》第 102 卷:49 http://www.pubmedcentral.nih.gov/picrender.fcgi?artid=1266160&blobtype=pdf 於 2008 年 4 月獲得。
- ↑ Cáceres, J 等人。 “可變剪接:多種控制機制及其在人類疾病中的參與”。《遺傳學趨勢》第 18 卷,第 4 期,2002 年 4 月 1 日,第 186-193 頁。 http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6TCY-45FYM7X-F&_user=47004&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_acct=C000005018&_version=1&_urlVersion=0&_userid=47004&md5=eea15989e03f8b963bdc33384a4ef93b
- ↑ “從 DNA 到 RNA”。《細胞的分子生物學》。 http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.figgrp.1020 於 2008 年 4 月獲得
- ↑ http://en.wikipedia.org/wiki/Proteomics
- ↑ http://en.wikipedia.org/wiki/Mass_spec
- ↑ “透過質譜法鑑定蛋白質”。美國能源部研究新聞 http://www.eurekalert.org/features/doe/2001-06/drnl-pib061902.php 於 2008 年 4 月獲得
- ↑ Lu, B 等人。 “透過結合使用 LTQ-Orbitrap 高質量精度進行鳥槍法蛋白質組學的 MS 和 MS/MS 資訊來提高蛋白質鑑定靈敏度”。《分析化學》,第 80 卷(第 6 期),2018-2025,2008 年。Datahttp://pubs.acs.org/cgi-bin/article.cgi/ancham/2008/80/i06/html/ac701697w.html 於 2008 年 4 月獲得
- ↑ Nesvizhskii, A 等人。 “鳥槍法蛋白質組學資料的解釋:蛋白質推斷問題”。《分子與細胞蛋白質組學》4:1419-1440,2005 年。 http://www.mcponline.org/cgi/content/full/4/10/1419 於 2005 年 4 月獲得
- ↑ Dralyuk, I 等人。可變剪接資料庫。 http://hazelton.lbl.gov/~teplitski/alt/ 於 2008 年 4 月獲得
- ↑ hanaraj T.A.、Stamm S.、Clark F.、Riethoven J.J.M、Le Texier V. 和 Muilu J. ASD:可變剪接資料庫。核酸研究。2004 年 32:D64-D69。 http://www.ebi.ac.uk/asd/ 於 2008 年 4 月獲得
- ↑ Bonizzoni, P 等人。 “可變剪接預測的計算方法”。《功能基因組學與蛋白質組學簡報》。第 5 卷(第 1 期),第 46-51 頁。 http://bfgp.oxfordjournals.org/cgi/content/full/5/1/46 於 2008 年 4 月獲得
- ↑ Black, D 等人。 “來自可變剪接的蛋白質多樣性——生物資訊學和後基因組生物學的挑戰”。《細胞》。第 103 卷,第 3 期,2000 年 10 月 27 日,第 367-370 頁。 http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6WSN-41T1FBT-2&_user=47004&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_acct=C000005018&_version=1&_urlVersion=0&_userid=47004&md5=d3b999a5147780e759f2e9aa7b8c47e7 於 2008 年 4 月獲得