
結構生物化學/衰老與代謝控制分析
衰老是指機體(如細胞)在其一生中積累了過量的損傷。由此產生的損傷最終會影響機體的整體生存和狀態。隨著機體衰老,“其輸出的退化導致功能下降和死亡,這是衰老的結果”(1)。這種輸出的退化與細胞的代謝歷史有關,而代謝歷史會影響細胞的功能。
衰老的概念導致人們普遍認為,衰老是由於機體在其一生中積累的損傷導致其無法保護、維持和修復自身。科學家正在進行實驗,試圖確定與衰老相關的哪些損傷會導致機體功能喪失。這很困難,因為與損傷相關的許多因素在不同模型和個體之間可能會有所不同,例如損傷程度、組織型別、年齡,以及所研究的機體型別。首先,機體要生存需要做些什麼?首先,機體必須能夠找到食物、住所,並抵禦感染或掠食者。機體必須能夠避免死亡。即使消除了這些因素,死亡和功能喪失也會隨著年齡的增長而發生。然而,並沒有進化出導致死亡的基因。衰老的機體被認為會降低個體對下一代的遺傳貢獻。換句話說,這是不利的。
在 Murphy 的論文“衰老的控制理論”中,他指出“基因組及其表達方式限制了死亡率和壽命”。然而,這種觀點仍然存在問題,因為所有機體的壽命各不相同,即使它們的基因非常相似。可以得出結論,導致死亡的年齡的確是遺傳決定、環境變化以及機體一生中發生的各種事件的綜合結果,而這些因素會對其產生影響。由於一生中低劑量暴露於紫外線輻射和伽馬射線輻射等外部因素,機體執行穩態機制的能力開始下降,衰老變得明顯。人們還提出,衰老反映了細胞衰老,即細胞自我複製和生長的能力不可逆轉地停止。
一個障礙是突變和環境干預,這些干預阻礙了許多功能,這使得很難確定衰老的原因。如前所述,機體死亡的原因可能有很多。重要的是要看到,必須考察所有可能的生化和生理實體,並獨立觀察每個實體。這將有助於縮小導致衰老的原因範圍。
層次結構的最高級別是機體的功能和一組參與相互作用以及與周圍環境相互作用的生理系統。每個系統都以某種方式透過輸入和輸出與其他系統以及環境相互作用。

在上圖中,為了簡便起見,相互作用的生理系統被顯示為兩個獨立的系統。它們受到來自環境和來自其他系統的輸入的影響。例如,隨著機體衰老,其各個系統的輸出隨著時間的推移而減少,機體的死亡率也會增加。對機體的損傷也會導致功能障礙,從而導致系統輸出不當。
一般來說,機體內的系統通常會隨著衰老而衰退。然而,很明顯,不同物種的死亡率並不相同。一種理論認為,自然選擇可能會影響機體的衰老過程。倖存下來的機體世代可以表現出可以透過進化保留的特徵。這暗示著不同機體(從果蠅、小鼠、酵母和蠕蟲)可能存在類似的衰老過程。然而,生物年齡並不可靠,因為這在不同物種之間也會有所不同。衰老的其他方面可能是器官特異性的,並且會表現為皮膚衰老的出現。對於一個獨立的系統,很難確定是缺陷輸入的影響還是該系統的內在損傷導致了細胞功能障礙。所有上述原因解釋了為什麼科學家難以確定是所有生理系統的衰退還是僅僅一個系統的衰退導致了機體衰老並最終死亡。
細胞會發生許多變化,但影響生理系統功能輸出的變化會影響衰老。系統的功能障礙是由於細胞數量及其輸出的減少造成的。根據 Murphy 的說法,“細胞的變化是由其代謝歷史引起的,並且是由於非特異性損傷以及訊號通路和基因表達的變化造成的。這些反過來會對細胞功能和細胞數量產生影響。”隨著機體衰老,組織會發生許多變化。衰老會導致細胞數量發生變化。細胞數量的減少會導致維持細胞的機制出現紊亂。
一些有絲分裂組織可以從其他分化細胞中補充失去的細胞。但隨著機體衰老,哺乳動物幹細胞補充失去細胞的效率會降低。為了確定生理系統是否因細胞數量的減少而受損,記錄系統受損之前和之後發生的細胞丟失或增益至關重要。細胞功能下降的一個例子包括神經元中的突觸傳遞和肌肉骨骼運動單位的收縮。在哺乳動物中,研究表明,隨著年齡的增長,由於突觸連線和傳導的減少,運動和神經功能會下降。
與限制飲食的動物有關的壽命成為機體對食物供應的反應,以及它們對環境中營養變化的反應。這些變化在細胞水平上透過能量啟用來衡量。餵養模式和覓食行為的變化也是其適應階段的指標。這些營養物質感應通路也為機體提供了時間來檢測和應對資源供應的變化。然而,由於這種壽命依賴於 daf-16,這意味著這些神經元的營養物質感應並非必需的。
現在我們來看看單個細胞,細胞功能障礙和死亡是細胞代謝歷史的屬性。這些屬性會影響系統功能,從而影響機體的死亡率。細胞的初始狀態取決於其基因組,其基因組會產生髮育史、機體中佔據的物理生態位以及影響基因組表達的表觀遺傳因素。
細胞的代謝歷史會導致 DNA 序列發生變化,而這些變化反過來會影響基因表達。這也會影響細胞在其一生中的增殖、功能障礙和死亡。
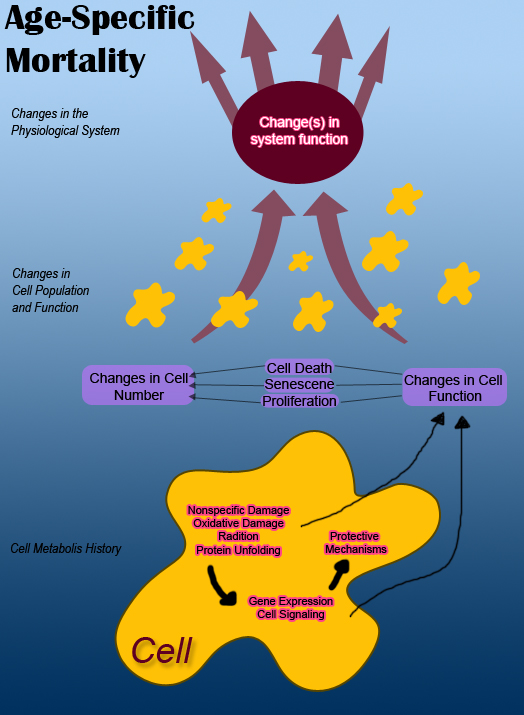

所有這些主題相互關聯,也與衰老過程相關。由於細胞數量及其功能的變化,系統會發生功能障礙。細胞數量和功能的變化是由細胞的代謝歷史引起的,主要涉及非特異性損傷和基因表達的變化。這些代謝歷史變化的結果導致細胞數量和功能的變化。影響衰老的細胞代謝歷史的重要因素和貢獻因素包括非特異性損傷以及基因表達和訊號通路的變化。
非特異性損傷
[edit | edit source]對生物體造成的不同型別的非特異性損傷會導致細胞功能喪失和功能障礙。一些損傷包括氧化損傷、輻射或化學反應。隨著時間的推移,非特異性損傷最終會損傷和阻礙分子。細胞盡力生存並保護自己免受損傷。在能夠修復錯誤摺疊的蛋白質的情況下,細胞會啟動損傷修復,但諸如 DNA 突變的修復之類的損傷無法解決。這種也是不可逆的非特異性損傷會嚴重損害細胞及其功能。隨之而來的問題是,對細胞造成的損傷是否會對系統的輸出產生足夠的影響以導致衰老。
基因表達和細胞訊號傳導
[edit | edit source]關於非特異性損傷,它會影響細胞的基因表達和細胞訊號傳導。受非特異性損傷影響的基因表達也會影響其對系統輸出的影響,從而影響正常衰老過程中的死亡率。有證據表明,在正常衰老過程中會發生大量的基因表達變化,但它們對衰老過程的貢獻會因系統而異,總體而言也會因生物體而異。
代謝控制分析與衰老
[edit | edit source]代謝控制分析 (MCA) - 一種實驗方法,用於瞭解控制在代謝途徑和網路中是如何分佈的。
為了確定與衰老過程相關的因素,必須對所分析的系統和觀察到的可測量變數進行設定限制。一旦這些因素被定義和限制,就可以測試和操縱所討論的生理系統,以便可以看到該變數與其控制的步驟之間的關係。在 MCA 實驗期間,該變數的控制程度也將被確定。透過使用 MCA,已經開發出強大的數學形式化方法來觀察整個系統,以及以下內容:定量確定控制程度、揭示所有控制部位以及控制步驟的影響。雖然已經表明,生化途徑和機制的極端和較大變化會增加或減少壽命,但沒有足夠的證據表明它們會影響正常衰老。與代謝途徑和衰老進行比較,MCA 可以告訴人們有關控制系統中特定過程的步驟的重要資訊。
MCA 在衰老中的應用
[edit | edit source]MCA 過程及其在系統中的應用需要以某種方式進行才能獲得結果。由於主題是生物體的衰老,因此需要與機體衰老相關的死亡率讀數。如前一節所述,可以對系統因素進行微小改變,並透過利用數學方法獲得可量化的結果。需要測量死亡率讀數以響應系統中所做的微小變化。請記住,因素的變化是由於認為它可能控制衰老。為了使一個因素影響衰老,它必須改變相應的死亡率和對衰老的貢獻。MCA 實驗是在從果蠅到小鼠的各種動物種群中進行的。然後可以測量死亡率讀數並進行比較,以檢視物種之間是否存在相似性。
在上圖中,死亡率讀數透過將其繪製在與衰老相關的變數(已增加和減少)上顯示。深藍色陰影區域表示變數在正常衰老過程中是如何改變的。該圖還顯示了正常衰老的不同情況。變數的影響可以降低或增加,這也影響死亡率。這是一個在 MCA 實驗後可以繪製的圖表示例,以展示一個人的發現和結果。主要思想是,如果選擇影響正常衰老的正確變數,死亡率曲線將具有可測量的斜率。
染色質在衰老中的作用
[edit | edit source]染色質 首先是在胚胎發生過程中在生物體中形成的。這種遺傳物質易於透過 DNA 甲基化和不同型別的組蛋白修飾進行修飾;這些變化不會影響 DNA 的核苷酸序列。這些變化在生物體一生中的積累導致隨機、隨機的結構。在一項對丹麥雙胞胎的實驗中,觀察到親屬之間的變異是染色質變化導致的遺傳轉移的結果。這支援了這樣一個觀點,即一種理論引用了對丹麥雙胞胎的實驗,並提出衰老是染色質結構變化積累的結果(2)。有幾種與染色質修飾相關的蛋白質。這些蛋白質直接影響轉錄調控,但也影響染色質中基因的結構和表達。
CDKN2a (p16INK4a)
[edit | edit source]CDKN2a 透過阻止 CDK4/CDK6 磷酸化視網膜母細胞瘤蛋白來調節視網膜母細胞瘤蛋白,視網膜母細胞瘤蛋白抑制腫瘤。CDKN2a 表達增加與細胞衰老和衰老相關,這與細胞壓力水平升高有關。由於這些關聯,已經建議使用這種蛋白質作為衰老的生物標誌物。在一項對小鼠 BubR1 半不足的實驗中,發現消除 CDKN2a 陽性細胞的受試者壽命延長,並且與年齡相關的退化減少。實驗還表明,在老年小鼠中去除 CDKN2a 陽性細胞改善了由於衰老引起的退化,並延長了這些影響。CDKN2a 表達受三胸 (TrxG) 和多梳 (PcG) 控制,它們分別開啟和關閉基因。
SIRT1 和 SIRT6
[edit | edit source]SIRT1 和 SIRT6 都是 NAD+ 依賴性賴氨酸脫乙醯基酶,在 DNA 修復和衰老中發揮作用。已在酵母中發現並對 SIRT1 和 SIRT6 的同源蛋白 Sir2 進行了實驗。這種蛋白質使 rRNA 和端粒沉默。如果沒有該基因的表達,那麼 rDNA 陣列將從基因組中去除以形成環狀 DNA,並且酵母的壽命會下降。類似地,SIRT1 和 SIRT6 會影響細胞對 DNA 修復和壓力水平的反應。這兩種蛋白質透過 DNA 修復過程和轉錄調控影響衰老過程。
當 DNA 發生氧化損傷時,SIRT1 會分散到所有修復位點,在那裡 H1K26 被脫乙醯化以增加 DNA 修復。然而,當 SIRT1 被招募到基因組的受損區域時,需要共濟失調毛細血管擴張突變途徑啟用,它會脫乙醯化 Nijmegen 斷裂綜合徵蛋白 (NBS1),NBS1 控制 DNA 修復途徑,但也控制損傷依賴性轉錄失調。因此,雖然 SIRT1 維持 DNA,但它在衰老中也起著至關重要的作用。SIRT1 表達增加不會延長生物體的壽命,但會增加其腫瘤抵抗力,改善其代謝,並預防糖尿病。SIRT1 還起著刺激非組蛋白的作用,例如 p53 (腫瘤抑制基因)、FOXO(轉錄框)和 Ku70(DNA 修復因子)。
然而,SIRT6 的過度表達確實會增加小鼠的壽命(尤其是在雄性小鼠中)。SIRT6 是一種調節蛋白,透過附著在活化 B 細胞核因子 κ 輕鏈增強子(NF-kB)的活性位點並抑制其轉錄來影響其依賴性炎症反應。因此,SIRT6 表達降低會導致 NF-kB 反應增加。缺乏 SIRT6 的人類更容易受到攻擊 DNA 的藥物的攻擊。SIRT6 還參與同源重組,調節對 DNA 損傷的反應,並透過增加端粒功能障礙來促進衰老。
總體而言,MCA 方法的開發極大地提高了我們對影響和影響正常衰老過程的因素的理解。MCA 方法應與測試衰老的實驗模型一起應用,因為它將使我們能夠解決可能影響衰老的因素。
- Murphy, Michael P. and Partridge, Linda. “Toward a Control Theory of Aging.” ["http://www.ncbi.nlm.nih.gov/pubmed/18318658"], 'Annual Review of Biochemistry', 2008.
- http://www.annualreviews.org/doi/pdf/10.1146/annurev.biochem.77.061206.171059
- O'Sullivan, Roderick and Jan Karlseder. "The Great Unravelling: Chromatin as a Modulator of the Aging Process". ("http://www.sciencedirect.com/science/article/pii/S0968000412001132"), 'Trends in Biochemical Sciences', 1 November 2012 (Vol. 37, Issue 11, pp. 466-476)