
結構生物化學/冷適應酶
生活在永久寒冷環境中的生物體擁有在寒冷環境中高效發揮作用的酶。一些例子包括魚類,它們也進化出抗凍蛋白作為一種適應方式,以及一些必須生活在寒冷地區的原核生物。最近的發現表明,對這些酶的進一步研究實際上可能具有生物技術應用。最常提到的冷適應酶是來自鹽生偽單胞菌(AHA)的α-澱粉酶,一種原核生物。生物體在寒冷環境中茁壯成長的能力來自於其合成冷適應酶的能力。在寒冷環境中繁榮的生物被稱為嗜冷菌。這些酶已經進化出一系列結構特徵,使其具有高靈活性,尤其是在活性位點周圍,低活化焓,低底物親和力和低溫下的高比活性。冷適應酶的結構、功能和穩定性的研究在蛋白質摺疊和催化研究中處於起步階段,這是一個仍在發展中的領域。
不同的生物體以不同的方式進化,導致它們適應不同的適合自身的熱環境。極端環境下的熱適應特別難以適應,並且單個生物體所能耐受的範圍有限。嗜冷微生物,即適應寒冷的微生物,據說能夠在-20°C的雪和冰中進行代謝。一些嗜冷菌甚至可以在≤0°C下增殖,但受限於<30°C的溫度(2-5)。
由於世界上許多海洋、寒冷的高山地區、洞穴、高層大氣和極地地區都發現了大量的微生物,因此地球上很大一部分生物量,或某個區域的活生物體,是在這些寒冷溫度下在其特定區域產生的。來自生命三個域的生物:細菌、古菌和真核生物,也來自這些寒冷的環境。大多數冷適應酶被發現來自原核生物。為了生存,所有這些生物體都必須與其周圍環境處於熱平衡狀態。這來自於其細胞的所有組分都適當地適應了寒冷。為了適應寒冷,已經採用了潛在的機械多樣性和細胞特異性適應策略。從對這些生物體的研究中出現的一個普遍模式是,生活在永久寒冷環境中的生物體通常會進化出有助於它們在寒冷環境中發揮作用的酶。

一個將被進一步研究和解釋的冷適應酶的例子是來自鹽生偽單胞菌(AHA)的α-澱粉酶。這種酶是被研究最多的冷適應酶。
下面的阿倫尼烏斯方程描述了所有反應(包括酶促反應)的速率。
kcat = AKe-Ea/RT
kcat = 酶反應速率(隨著絕對溫度(T)的升高而增加,隨著活化能(Ea)的降低而降低)
A = 指前因子
K = 動態傳遞係數(通常假定為1)
R = 普適氣體常數(8.314 J mol−1 K−1)
根據該方程,由於0°–4°C的低溫,系統可用於克服障礙的動能不足。一些有助於補償緩慢代謝速率的策略包括:增加酶濃度的能量消耗策略,魚類和線蟲中同工酶的季節性表達,以及酶的進化,其中反應速率趨於變得與溫度無關,而是接近擴散控制。冷適應酶傾向於將其最佳活性溫度轉移到較低的溫度,同時穩定性降低。這些酶在降低基態和過渡態之間的活化自由能壘時表現出高反應速率,如下式所示
ΔG#=ΔH# -TΔS#
ΔG# = 活化自由能壘
ΔH# = 活化焓的變化
ΔS# = 活化熵的變化
T = 絕對溫度
冷適應酶的結構具有高度的靈活性,這使得它們在低至中等溫度下解摺疊。研究人員嘗試了許多方法來研究解摺疊和摺疊轉變,以確定它們解摺疊的方式並測量酶的動力學和構象穩定性。這些方法包括分光光度法、量熱法和電泳法。已經使用這些技術對多結構域蛋白質(如殼寡糖酶)進行了實驗。蛋白質有解摺疊的趨勢,並且這些過程是不可逆的。當大的多結構域蛋白質因熱而解摺疊時,這些蛋白質傾向於以動力學驅動的形式完成解摺疊過程。
-
 殼寡糖酶的結構
殼寡糖酶的結構

AHA是唯一一個具有可逆解摺疊的冷適應酶的例子。在20°C或更高的溫度下,AHA具有可逆解摺疊,這可以透過在初始熱變性後的第二次掃描中100%恢復ΔH cal.(解摺疊過程中吸收的總熱量)來證明。小分子量酶透過協同解摺疊過程解摺疊。協同解摺疊發生是因為其緊密堆積的體積導致結構元素之間相互作用的數量很少,這有助於它保持其自然狀態。如果協同解摺疊中數量有限的相互作用受到干擾,則由於相互作用的增加,可能會發生兩態解摺疊。
在胰腺豬α-澱粉酶(PPA)和更穩定的AHA突變體中,不會發生兩態解摺疊過程。相反,其他型別的摺疊可能來自離子相互作用的增加,導致摺疊過程中分子內差異的頻率發生。AHA突變體顯示了這種差異或發散的概念,導致熱失活速率與可逆程度直接相關。
在TUG-GE酶中,在3°C和12°C的溫度下,AHA可逆地解摺疊並顯示出兩個轉變。在較低尿素濃度下解摺疊的轉變屬於活性位點區域。活性位點是由結構的協同解摺疊形成的,顯示了蛋白質其他更穩定區域或結構域的獨立解摺疊。當解摺疊開始時,發現冷適應酶的底物結合區域是最靈活的區域,因為其高Km。對AHA的研究表明,活性位點區域的不穩定性是熱不穩定酶的關鍵,並且是檢查更廣泛的冷適應酶的重要概念。
酶失活或變性的過程用動力學穩定性表示。大多數冷適應酶在50°C的溫度下半衰期小於20分鐘,有些酶甚至在較低的溫度下變性。為了預測動力學穩定性,必須考慮摺疊或活性狀態與過渡狀態之間的自由能變化的大小,如下所示
F K¦↔ 〖TS〗^# k¦→ D
F = 摺疊酶
K = 平衡常數
K= 一級速率常數
D = 變性狀態
冷適應酶透過降低ΔG#來提高熱解摺疊速率,如下式所示
ΔG#= =RT ln K
冷適應酶的熱穩定性降低可能是由於摺疊形式的ΔH#較低。需要斷裂一定數量的一價相互作用才能達到過渡態(TS#),並且此數量的減少導致摺疊形式的數量減少。穀氨酸脫氫酶是一種熱穩定性低的酶的例子,這是過渡態無序性增加的直接結果。在解摺疊過程中,熱穩定酶熵降低的關鍵原因可能是非極性基團的水合作用。這是由水圍繞疏水側鏈形成有序結構引起的,從而降低了系統的熵。確定變性的活化引數對於發現更大範圍的冷適應酶及其熱穩定同源物至關重要。
透過將冷適應酶的X射線結構與來自中溫生物和嗜熱生物的蛋白質同源模型進行比較,已經發現X射線結構揭示了冷適應的結構基礎。酶的活性與穩定性與其生理環境密切相關。研究還表明,主要利用海洋生物來研究冷適應酶。已經確定了檸檬酸合酶的X射線結構,並將其與細菌、真核生物和古菌的其他酶進行了比較。由於缺乏晶體結構資訊,研究通常選擇密切相關的生物體。透過最大化比較資料集中的熱力學差異並最小化系統發育差異,可以彌補酶X射線結構的不足。
疏水殘基與溶劑水分子之間的疏水相互作用在促進冷適應蛋白質的結構靈活性和熱穩定性方面發揮著關鍵作用。
冷適應酶的氨基酸往往比中溫生物和嗜熱生物的同源物更小且疏水性更低。由於範德華力較弱、作用範圍短且對距離敏感,蛋白質內部疏水基團之間的距離將決定對穩定化的焓貢獻。因此,範德華力的減少和內部基團運動的增加將導致冷適應酶的不穩定。蛋白質內部的一個基團例子是異亮氨酸。由於其支鏈和大小,異亮氨酸可以有效地填充核心並穩定蛋白質。在冷適應的胰蛋白酶、檸檬酸合酶和AHA中,核心內異亮氨酸殘基的數量更少。
疏水相互作用在室溫下最強,因為疏水側鏈在水中的溶解度在20°C的最低溫度下達到最小。透過對31種可逆展開蛋白質的研究,發現約75%的蛋白質在室溫附近具有最大穩定性。這表明蛋白質核心中的疏水相互作用在增強蛋白質在低溫到中等溫度下的穩定性方面發揮著關鍵作用。
許多酶的X射線結構研究表明,冷適應酶的表面存在較高比例的疏水或非極性殘基。使用大規模建模和結構研究,發現了類似的結論,這些結論表明酶的可溶劑化表面或埋藏表面的平均分數在冷適應蛋白質中具有更高的疏水性。在異亮氨酸的例子中,X射線結構顯示,在冷適應酶中,異亮氨酸在亞基介面處的聚集不存在,而在來自超嗜熱菌的同源物中則存在緊密堆積的疏水簇。
研究表明,疏水錶面殘基會由於水分子熵的降低而導致蛋白質結構不穩定,因為這些水分子會在非極性殘基周圍形成籠狀結構。研究發現,在較低溫度下,由於釋放的水分子遷移率降低,熵增實際上會減少,這表明冷適應酶可以從增加的表面疏水性中獲得靈活性和更大的耐受能力。
某些冷適應酶,如胰蛋白酶和β-內醯胺酶(55),表面電荷增加,主要是負電荷。對於某些酶,負電荷相當高,但在活性位點附近也存在一些正電荷。由於水在低溫下具有高粘度和高表面張力,破壞氫鍵網路的能量成本很高。這種能量成本可以透過與具有高介電常數的水分子相互作用的表面帶電或極性氨基酸來抵消。這將有助於適當的溶劑化並幫助維持酶的靈活性。透過更好的溶劑相互作用和正電荷,可以在低溫下提高靈活性。
-
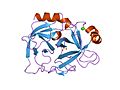 胰蛋白酶的結構
胰蛋白酶的結構

表面斑塊中酸性殘基的定位可能產生導致蛋白質結構整體不穩定的電荷-電荷排斥。有時會出現酸性殘基的電荷排斥可能在連線區產生高水平靈活性的情況。這是某些酶冷適應的主要結構特徵。其他研究表明,冷適應涉及溶劑可及和埋藏表面的平均分數減少。這涉及不僅來自古菌家族,也來自細菌家族的其他蛋白質,導致蛋白質表面親水性增加。透過增加熱穩定蛋白質的表面電荷,研究人員能夠將其與形成鹽橋網路的能力聯絡起來。這與冷適應酶中與水分子相互作用形成直接對比。已經發現,隨著離子相互作用隨著溫度降低而增強,其數量最小化,從而使冷適應蛋白質在低溫下保持靈活性。
Siddiqui KS, Cavicchioli R. 冷適應酶。生物化學年度評論。第75卷:403-33。卷出版日期2006年7月。