
結構生物化學/朊病毒蛋白錯誤摺疊和疾病
傳染性海綿狀腦病(TSE 或朊病毒病)是一組罕見的致命性神經退行性疾病,影響人類和其他哺乳動物。TSE 是蛋白質錯誤摺疊疾病,包括正常宿主朊病毒蛋白異常積累形式的聚集。TSE 的獨特之處在於它們具有傳染性。TSE 的特徵包括它們可以複製,能夠進行選擇性進化(可以進化出耐藥性),具有多種與體內獨特表型相關的感染因子菌株,並表現出強烈的種屬特異性;這是許多病毒和細菌病原體的相同特徵。

人類 TSE 疾病分為 3 類
- 散發型 - 最常見的 TSE 形式(例如克雅氏病)
- 遺傳型 - TSE 是朊病毒蛋白內的突變
- 獲得性 - TSE 是攝入或接種受 TSE 汙染的物質的結果。
-
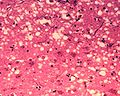 顯微鏡下的“孔洞”是朊病毒感染組織切片的典型特徵,導致組織形成“海綿狀”結構。
顯微鏡下的“孔洞”是朊病毒感染組織切片的典型特徵,導致組織形成“海綿狀”結構。

典型的感染因子使用核酸來傳播和增殖,在細菌和病毒中很明顯。朊病毒假說簡單地說,上述特定疾病完全由蛋白質引起,這一觀點在該理論提出時與當時的常識相悖。起初,人們認為朊病毒是其他型別感染的副作用。然而,這一理論站不住腳。自提出朊病毒誘發疾病的概念以來,實驗證據支援了這一理論,朊病毒複製的影響在實驗室中已經得到證實。 [1]
朊病毒及其傳染性最早是在 1937 年發現的,當時在進行病毒接種時,羊瘙癢病意外地傳染給了綿羊。在後來的實驗中,羊瘙癢病被有意地傳染給綿羊,然後傳染給小鼠,以確定該分子的性質。新幾內亞的食人行為是庫魯病感染人類的來源,1966 年證明該疾病可以傳染給猴子[1]。此時,科學界開始瞭解到朊病毒的傳染性不同於病毒或微生物感染。最近,牛海綿狀腦病和與食用受感染肉類相關的變異克雅氏病的爆發被證明是由朊病毒引起的。
從歷史上看,對朊病毒的研究首先確定了感染方式是新穎的,然後確定它是由於蛋白質鏈的錯誤摺疊造成的。第一個實驗使用紫外線和電離輻射破壞核酸,發現感染因子仍然存在[1]。然後,確定仍然具有感染性的最小分子量顆粒的重量約為蛋白質的重量。
後來,發現抗蛋白酶的朊病毒蛋白(PrP)濃度與感染性成正比。此外,使用破壞蛋白質結構的試劑,降低了 PrP 的感染性。發現 PrP 存在於正常運作的大腦中,證明該蛋白質既可以以正常形式存在,也可以以故障形式存在[1]。最近,研究表明,感染性和非感染性蛋白質可以混合,感染因子會在非感染性分子中傳播。這表明蛋白質摺疊錯誤可以無限期地傳播。
TSE 是一種蛋白質錯誤摺疊疾病,疾病發生是由於宿主朊病毒蛋白 (PrP) 的構象發生改變。PrP 是一種哺乳動物糖蛋白,長 209 個氨基酸。當 PrP 變成 TSE 時,在一個稱為發病機制的過程中,PrP 的一種抗蛋白酶敏感形式 (PrP-sen) 會重新摺疊成 PrP-res(一種抗蛋白酶的朊病毒蛋白形式)。PrP-res 和 PrP-sen 具有相同的初級序列,但二級結構不同,PrP-sen 具有更多 α-螺旋。(換句話說,PrP-res 和 PrP-sen 是同種型)PrP-res 是 TSE 的主要成分。因此,研究 PrP-sen 如何轉化為 PrP-res 非常重要,以便抑制 PrP-res 形式的形成。

目前對 PrPc 到 PrPSc 轉化的瞭解可以概括為一個兩步機制
1) 前體“種子”PrPSc 的有序聚集體與 PrPc 結合。
2) PrPc 經歷某種構象變化,導致其傳播成更多 PrPSc。這一步的機制在很大程度上是一個謎。轉換後的 PrPSc 被新增到聚合物中,該聚合物最終會斷裂並導致更多 PrPc 被轉換。這種斷裂被認為是反應的限速步驟。 [1]
2001 年,開發了一種在實驗室複製朊病毒的方法。這種方法被稱為蛋白質錯誤摺疊迴圈放大 (PMCA),體外產生模擬體內正常複製的朊病毒。該過程依賴於朊病毒是自催化的,在正確環境下可以無限期地複製。這種技術被證明對研究朊病毒和檢驗朊病毒假說非常寶貴。 [1]
科學家們目前還不清楚輔助因子在朊病毒複製過程中的重要性。雖然現在已經瞭解到有一些外部因素會影響朊病毒複製的成功率和速度,但這些輔助因子尚未完全確定,並代表著未來研究的一個領域。
關於輔助因子在朊病毒複製中作用的證據來自對動物物件的各種研究。在一項對轉基因小鼠進行的研究中,發現存在某種影響朊病毒蛋白表達的因子。這被稱為“蛋白 X”,儘管該因子的身份從未被確定為一種蛋白質。 [2]. 在一項對倉鼠 PrPc 進行的獨立研究中。當分離和純化時,倉鼠 PrPc 在與 PrPSc 混合時無法轉化。當向樣品中加入腦勻漿時,朊病毒轉化發生,表明勻漿中存在某種物質有助於轉化,無論是作為催化劑還是作為組成部分。進一步測試表明,倉鼠中的 RNA 充當朊病毒複製的催化劑,但在小鼠中則不然。具體是如何做到這一點尚不清楚,但有可能它有助於穩定所產生的 PrPSc 的構象。它並非對所有哺乳動物都有效這一事實導致了多種輔助因子的可能性,或者物種/器官特異性輔助因子。
雖然輔助因子的重要性尚未完全瞭解,但輔助因子可能影響朊病毒轉化的方式可分為 5 類。
| 遺傳資訊 | 可能存在某些輔助因子透過幫助確定這些蛋白質的摺疊方式來促進朊病毒複製。一項支援這一觀點的研究是在小鼠中進行的,透過將朊病毒注射到不同型別的細胞中。在每個細胞中,朊病毒複製導致不同的朊病毒菌株,這可能是每個細胞型別中不同輔助因子導致的結果。 |
| 催化 | 某些輔助因子可能透過附著在 PrPc 上並部分展開它來催化 PrPc 到 PrPSc 的轉化。這將使 PrPSc 更容易在蛋白質上誘導錯誤摺疊模式,導致轉化。這種型別的輔助因子在實驗室中進行了體外研究。 |
| 構象穩定 | 一些輔助因子可能有助於穩定新的朊病毒構象。這類輔助因子的例子包括核酸、蛋白質、金屬離子和其他物質。其中許多是帶電物質,可以與朊病毒結合並幫助形成更緊湊的結構。 |
| 片段化 | PrPSc 聚合物通常會片段化,這極大地加速了 PrPc 到 PrPSc 的轉化。一些輔助因子可能有助於這一片段化過程,這會產生新的種子,對複製過程至關重要。在酵母中發現了一個例子,它依賴於蛋白質 104 (Hsp104)。去除這種蛋白質會阻止朊病毒複製的發生。 |
| 生物穩定 | 為了成功地進行朊病毒複製,朊病毒必須首先在生物介質中存活下來。某些生物抗性,例如可以進行吞噬作用的小膠質細胞,可以抑制它們的傳播。一種降低小膠質細胞破壞這些朊病毒能力的輔助因子將允許它們複製,並且可能被證明是必不可少的。 [1] |
PrP-sen 中氨基酸 108-189 內的一級和二級結構成分被證明對轉化很重要,如核磁共振結構和體內研究所確定。PrP-sen 的關鍵殘基 (108-189) 包括大部分摺疊域,包括 β 摺疊、α 螺旋,最重要的是環和轉角。轉角和環的輕微變化特別值得注意,因為已經確定環參與 PrP-sen 和 PrP-res 之間相互作用。
雖然對 PrP-sen 三級結構有廣泛的瞭解,但對 PrP-res 卻不能這麼說。關於轉化為 PrP-res 的機制途徑缺乏廣泛知識的部分原因是 PrP-res 結構未知,因為它尚未被充分純化以進行高解析度結構研究。由於無法使用高解析度技術,確定 PrP-res 結構的重點一直侷限於使用低解析度技術,例如電子顯微鏡來確定 PrP-res 的超微結構和二級結構。
在酵母中發現了一種有趣的朊病毒案例。雖然它們通常在哺乳動物中觀察到,但發現酵母中的一種特定蛋白質 (Ure2) 以典型朊病毒的方式表現和繁殖。然而,它並沒有殺死宿主細胞,而是透過誘導其他蛋白質的構象變化而繁殖,並透過細胞分裂遺傳。這種特殊情況導致了在真菌中發現其他朊病毒,這些朊病毒也表現出非致死性行為。 [1]
{kind=link}
正如在酵母朊病毒的例子中發現的那樣,這些錯誤摺疊的蛋白質並非天生具有惡意,也不對有機生命有害。在西南大學科學家進行的一項研究中,發現體內某些朊病毒樣蛋白質可能有助於免疫系統。這些線粒體抗病毒訊號蛋白 (MAVS) 被發現存在於線粒體膜中,並積極防禦病毒感染。研究發現,在受到病毒感染威脅的情況下,MAVS 蛋白會錯誤摺疊並在線粒體表面聚集,有效地保護該細胞器免受攻擊。這種有意的錯誤摺疊和聚集很有趣,因為它並不反映朊病毒在克雅氏病和牛海綿狀腦病等情況下引起的惡意和不可控的錯誤摺疊。雖然初始病毒檢測對系統的順序啟用可能很複雜,但 MAVS 蛋白的聚集實際上發生得很快。 [4]
PrP-res 自身放大的方式與澱粉樣蛋白纖維透過從前體蛋白庫中種子聚合而生長的方式高度相似。因此,澱粉樣蛋白纖維為 PrP-res 生長機制的假設提供了啟示。
澱粉樣蛋白是纖維狀蛋白質聚集體,具有特定的結構特徵。澱粉樣蛋白纖維的結構具有特徵性的交叉β摺疊模式,其中β摺疊排列形成片層,這些片層垂直於纖維軸,而鏈之間的氫鍵平行於纖維軸。雖然這種交叉β四級結構在澱粉樣蛋白中是已知的,但尚未確定 PrP 的這種範圍內的詳細結構,PrP 是迄今為止更大的蛋白質。
與澱粉樣蛋白纖維一樣,PrP 可以被誘導形成澱粉樣蛋白樣纖維。一個特別重要的模型是重組肽 PrP23-144。 (重組肽是從其相應的重組 DNA 編碼的肽)。PrP23-144 被證明具有高度的構象可塑性 (能夠以各種可能的方式改變構象),並且可以被誘導形成多種纖維氨基酸。該模型中 PrP23-144 可以被區分成多種構象可能性,這表明 PrP 同種型轉換高度依賴於 PrP-sen 到 PrP-res 之間的結構相容性 (例如,前面提到的轉角和環的變化)。溶劑的選擇也起著作用,因為某些結構元素會受到靜電或親水效應的影響。
治療
[edit | edit source]由於科學家對朊病毒的瞭解有限,目前還沒有針對診斷為朊病毒相關疾病的患者的完全可靠的治療方法。朊病毒清除的一種可能性是透過小膠質細胞和吞噬作用。也發現地衣具有分解朊病毒的能力。[5] 儘管有很多可能的科研方向,但對於朊病毒相關疾病患者來說,還沒有臨床解決方案。
檢測
[edit | edit source]
在過去十年中,研究已經幫助確定了血液中的 PrPSc。直到現在,還沒有可靠的方法來識別腦組織中的這些錯誤摺疊的蛋白質。科學家採用一種稱為 SOFIA (環繞光纖免疫測定) 的新方法,能夠以高精度 (十億分之一) 識別腦組織中朊病毒的存在。首先,使用抗體放大並用熒游標記含有 PrPSc 的樣品。然後將其放入一個裝置中,該裝置用可以檢測發射光的環繞光纖包圍樣品。使用雷射激發染料,探測器拾取來自樣品的發射。
這種方法在以前被認為健康的綿羊身上進行了測試,但最終患上了朊病毒病。分析了它們在出現症狀前的血液樣本,發現可以非常早地檢測到 PrPSc 的存在。雖然這種方法不能治癒或消除任何問題,但它可以用來隔離並極大地降低這些朊病毒傳播的風險。[6]
- ↑ a b c d e f g h Soto, Claudio "朊病毒假說:爭議的終結?", '[PubMed]', 2010. 2012 年 11 月 20 日檢索。
- ↑ Telling GC, et al. “在表達人類和嵌合 PrP 轉基因的小鼠中朊病毒的傳播暗示了細胞 PrP 與另一種蛋白質的相互作用。”, ‘[PubMed]’, 2012 年 6 月 27 日。 2012 年 11 月 20 日檢索。
- ↑ Weissmann C. "關於哺乳動物朊病毒株的想法", '[PubMed]', 2009. 2012 年 11 月 20 日檢索。
- ↑ 侯法建等。 "MAVS 形成功能性朊病毒樣聚集體以啟用和傳播抗病毒先天免疫反應", '[Cell]', 2011. 2012 年 11 月 20 日檢索。
- ↑ Johnson, Christopher J. 等 "地衣中絲氨酸蛋白酶對疾病相關朊病毒蛋白的降解", '[PLOS]', 2011. 2012 年 11 月 20 日檢索。
- ↑ Rubenstein, R., Chang, B., Gray, P., Piltch, M., Bulgin, M.S., Sorensen-Melson, S. & Miller, M.W "在血液中檢測朊病毒", '[Microbiology Today]', 2010. 2012 年 11 月 20 日檢索。
參考文獻
[edit | edit source]- Moore, R (2009) 朊病毒蛋白錯誤摺疊和疾病